Dogma periférico: ¿de qué mensaje me están hablando?
Pablo Esteban Rodríguez
Instituto de Investigaciones Gino Germani, Facultad de Ciencias Sociales, Universidad de Buenos Aires. Buenos Aires, Argentina
Recibido: 26/05/2015 - Aceptado: 31/07/2015
Resumen
El artículo traza una analogía entre la teoría de la comunicación social y la descripción corriente sobre la relación entre DNA, RNA y proteínas, basándose en su referencia común a la teoría de la información y a su esquema más simple de la comunicación interpersonal e intermolecular: emisor, mensaje, receptor, y junto a ellos el código, el canal y el contexto (modelo de Jakobson). En ambos casos se observa un desplazamiento desde la importancia dada a la posición de emisión y al contenido del mensaje hacia la posición de recepción, lo que en términos de la biología molecular obligaría a proponer la elaboración de un Dogma Periférico que postula la existencia de una semiosis genética, en contraposición de un Dogma Central basado en la secuencia genética.
Palabras clave: comunicación, información, recepción.
Peripheral dogma: ¿what message are you talking about?
Summary
The article traces an analogy between the theory of the social communication and the current description about the relation DNA-RNA-proteins; an analogy based on its shared reference to the theory of the information and to the simplest scheme of the interpersonal and intermolecular communication: sender, message, receiver, and together with them the code, the channel and the context (Jakobson’s model). In both cases it can be observed a displacement from the centrality given to the position of the sender and to the content of the message towards the position of the receiver, which in terms of the molecular biology would force to propose the elaboration of a Peripheral Dogma. This new Dogma proposes the existence of a genetic semiosis, in contrast with the Central Dogma based on the genetic sequence.
Keywords: communication, information, reception.
En las teorías de la comunicación basadas en la teoría matemática de la información es tradicional definir a un emisor, un mensaje y un receptor. El reconocido lingüista y fonólogo Roman Jakobson tomó el clásico modelo de Claude Shannon [1] y lo amplió incluyendo al código, al canal y al contacto [2]. Reemplazando al contacto por el canal, al destinador (aquella instancia que motiva al sujeto a cumplir su objetivo, una fuerza que mueve al sujeto a ejercer una función) por el emisor y al destinatario por el receptor, según la acepción corriente en las disciplinas de la comunicación social, el esquema es el siguiente:
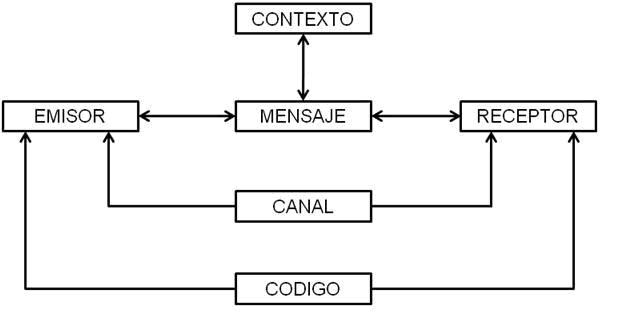
Figura 1: Modelo ampliado de la comunicación. (http://lenguayliteraturacastellana2.blogspot.com.ar/p/las-funciones-del-lenguaje.html)
En el campo de la comunicación social estos modelos fueron sometidos a constante crítica, en especial a partir de la década del ‘70. Se los acusó de presentar la acción comunicativa como lineal, concentrada en el emisor o, en el mejor de los casos, en el mensaje y en el código. En el ámbito de lo social, esto equivalía a desestimar la función del receptor en tanto creador de mensajes, o sea, emisor (Figura 2). Para estas posturas, no habría una flecha que va de un polo a otro sino una codeterminación de las instancias de emisión y recepción, lo que hace que los mensajes se transformen constantemente en función de otras instancias que habían sido dejadas de lado, como el contexto. Así, las teorías lingüísticas duras dieron paso primero a las teorías de la enunciación (quién dice qué en qué contexto y con cuál historia) y luego a las teorías de los discursos sociales, que estudian la emisión y la recepción en un pie de igualdad y afirman que aquella flecha emisor-receptor corre en varios sentidos. La semiosis es cualquier forma de actividad, conducta o proceso que involucre signos, incluyendo la creación de un significado. Es un proceso que se desarrolla, según el creador de la lingüística moderna, Ferdinand de Saussure [3], en la mente del intérprete; se inicia con la percepción del signo y finaliza con la presencia en su mente del objeto del signo. La semiosis social, esto es, la producción ya no individual sino social de sentido, es un conjunto de estados que el análisis identifica como productos y a través de los cuales se analizan los procesos; un recorte, una fotografía de un movimiento incesante [4]. Ya en la década del ’80 comenzaron a desarrollarse las teorías de la recepción [5] que retomaban las investigaciones realizadas por la Escuela de Birmingham, una de las principales fuentes de la teoría de la comunicación en el siglo XX.

Figura 2: l receptor en tanto emisor [3].
¿Las moléculas se comunican?
¿Qué tiene que ver todo esto con las proteínas? La cibernética, definida como ciencia que estudia la comunicación y el control en animales, hombres y máquinas [6], reunió una serie de investigaciones de campos muy diversos, con preeminencia de la ingeniería en telecomunicaciones, la matemática y la lógica. La teoría matemática de la información fue integrada allí y uno de los primeros resultados exitosos fue la computadora tal como la conocemos hoy, concebida explícitamente como un cerebro artificial. El proyecto de la cibernética era extender los avances de la informática, en especial estos esquemas de la comunicación, a diferentes ciencias y disciplinas [6,7].
Max Delbrück, físico que con los años obtendría el Premio Nobel en 1969 por su investigación con virus bacteriófagos junto a Salvador Luria y Alfred Hershey, asistió en 1946 a las conferencias fundacionales de la cibernética [8] y salió espantado por la poca seriedad de lo que había escuchado. Sin embargo, pocos años después se recurría masivamente a las metáforas informáticas y comunicacionales para explicar por qué los fenómenos específicos de la vida consisten en particulares arreglos moleculares. Así surgió el famoso Dogma Central de la biología molecular, enunciado por J.Crick [9], en el cual se postuló que la información va de modo lineal del DNA (emisor) a las proteínas (receptor) llevando un mensaje a través de un canal (RNA“mensajero” o “ de transferencia”), con un código aceptado (el código genético).
Pocos años después, en 1970, el mismo Crick incluyó dentro del Dogma Central la transcripción reversa de RNA a DNA, en parte como respuesta a un artículo de Temin y Mitzutani en el que afirmaban haber descubierto una DNA polimerasa dependiente de RNA, lo que luego se llamaría transcriptasa reversa [10,11,12]. Se puede entender este agregado como un intento de modular el Dogma Central, y a él se agregaron otras variantes. Sin embargo, en lo esencial, y más aún a partir del hallazgo del DNA recombinante y de su capacidad para inducir modificaciones genéticas en bacterias [13,14], el Dogma Central mantuvo su vigencia por mucho tiempo. El mismo Crick, por ejemplo, en un artículo conjunto con Orgel, volvió a salir al paso de posibles objeciones que pusieran en riesgo su validez cuando habló del famoso “junk DNA”, para referirse al DNA que no es transcripto en RNA y parece no cumplir ninguna función según el Dogma [15].
Dejando de lado la perplejidad que puede causar que la biología acepte nombrar a uno de sus fundamentos con el término netamente religioso de “dogma” [12], es sorprendente encontrarse con que las moléculas “se comunican” según un esquema que mezcla los modelos unilineales derivados de la información con analogías con las computadoras; más aún cuando en la actualidad las ciencias biológicas emplean cada vez más a la bioinformática. De esa mezcla se deriva que los mensajes son en lo esencial instrucciones y que puedan comprenderse como algoritmos, pues podrían ser cualquier otra cosa. En este sentido, se puede decir que el complejo DNA-proteínas fue primero entendido como una computadora y que más tarde ella misma, como devolución epistemológica de favores, se transformó en herramienta de análisis del funcionamiento de ese complejo.
¿Las moléculas interpretan?
Según el Dogma Central, las secuencias genéticas implicadas en los procesos de transcripción y traducción generan las estructuras, que serán las proteínas, que tienen innumerables funciones. Esto corresponde con los modelos de la comunicación de la primera época. Sin embargo, como toda ciencia de la comunicación del siglo XX, la biología molecular ahora también discute este esquema a favor de una consideración más atenta al contexto y a la posición de recepción. Ya en los años ’70 y ’80, con los ejemplos ya citados de la transcripción reversa de RNA a DNA y el “junk DNA” desarrollados por el propio Crick, la unidireccionalidad del Dogma Central se enfrentaba con algunos problemas.
En la actualidad se ha incorporado todo aquello que “no entra” en el Dogma original. Primero, el comportamiento de los priones, partículas patógenas de naturaleza proteica y con ausencia de ácidos nucleicos (responsable entre otras cosas de la conocida enfermedad llamada “de la vaca loca” o enfermedad neurológica de Creutzfeldt-Jakob), mostraría que “la información fluye de las proteínas hacia el genoma vía la asimilación de la variación epigenética” [16]. Segundo, el metabolismo celular “debe ser incluido en el flujo de información”, que ya no se produciría en sentido único, pues “la red bioquímica tiene su propia dirección autónoma que limita la ejecución de las instrucciones del DNA” y así es posible afirmar que hay un proceso de feedback entre dicho metabolismo y el DNA [17]. Tercero, en las últimas dos décadas, gracias a los avances en los procesos de secuenciación del RNA, se verifica que esta molécula es “un modulador de la expresión genética tanto pre como pos-transcripción, un componente de redes computacionales, una entidad alostérica, una molécula de señales y un agente de la inmunidad adquirida” [18].
Finalmente, el papel de las proteínas en el proceso de duplicación del DNA e incluso en el arreglo de su disposición espacial (histonas), las redes de comunicación intercelular, los mecanismos de feedback en los procesos celulares que permiten hablar de una “regulación epigenética”, entre otros procesos hallados en las últimas décadas, constituyen ejemplos de los límites del Dogma Central [12]. Otras investigaciones señalan incluso que sería posible encontrar procesos de traducción reversa, esto es, de proteínas a RNA [19].
Esto querría decir que las moléculas intervinientes en los procesos genéticos no sólo obedecen a una instrucción (en caso de que la haya), sino que implican al contexto y de ese modo “hacen” algo con aquello que reciben, lo interpretan. Ni la emisión es solamente acción ni la recepción supone únicamente pasividad. Este modo de comprender la comunicación biomolecular según una extraña “teoría de los discursos sociales” moleculares diría, pues, que hay secuencias, que de allí se pasa a las estructuras pero que, antes de pasar a las funciones, habría justamente actividad e interacciones. Si las estructuras (proteínas) cambian su conformación e integran al contexto (ambiente) dentro de su actividad, y a la vez, como se sabe, también regulan en cierto sentido la propia expresión de la secuencia (determinando en qué lugar se cortan las largas tiras de DNA), se deriva entonces que las propias secuencias podrían no ser completamente estables, y de hecho se sabe que los genomas reaccionan al entorno. Por lo tanto, cabría pensar que la genética, entendida como el estudio de la transmisión de la herencia o información codificada en los genes, no tendría por qué concentrarse sólo en la zona que lleva escrito el código, es decir, únicamente en los procesos de transcripción y traducción. Los ácidos nucleicos fueron identificados en los cromosomas por Johannes Miescher en 1869 y hubo que esperar más de 70 años hasta que Oswald Avery, Colin Mc Leod y Maclin Mc Carthy demostraran que el DNA era de hecho el material genético [20].
¿Las moléculas hablan?
Claude Shannon, el autor de la teoría matemática de la información, era ingeniero en telecomunicaciones. Estudiaba la manera más eficaz de establecer una comunicación telefónica en tiempos (década del ’40 del siglo XX) donde el ruido de fritura era importante. Las teorías de la comunicación, las alineadas y las críticas, siempre consideraron a seres hablantes, que producían significaciones, que generaban sentido. Al observar un proceso corriente de interacción entre DNA, RNA y proteínas se ven muchas cosas, entre ellas procesos de interacción física, pero no se ve a nadie diciéndole nada a nadie.
En este sentido, pareciera que las derivas del Dogma Central en los últimos 40 años replican las de las teorías de la comunicación en el mismo periodo. Fue Jakobson quien a principio de los ’70, luego de haber adaptado la teoría matemática de la información a un esquema general de la comunicación, confirmó la analogía entre secuencia genética y secuencia verbal [21, 22], dándole un sesgo eminentemente lingüístico al punto de vista informacional de la genética.
Sin embargo, la importancia adquirida por la teoría de los discursos sociales, y luego por las teorías de la recepción de los medios masivos de comunicación, mostró los límites del esquema de la comunicación derivado de la lingüística tradicional (Saussure, Jakobson). Se enfatizó entonces el papel del contexto y la reversibilidad entre emisores y receptores respecto del centro del esquema: emisor-mensaje-receptor-código. Y esto mismo parecen demostrar los “descubrimientos post-Dogma Central” [12], particularmente en lo que tiene que ver con la actividad del RNA y las proteínas y la regulación epigenética a través de otros actores intervinientes, como la célula y su metabolismo. En este sentido, cabría sostener la hipótesis, a explorar en trabajos futuros, de la existencia de una semiosis genética, más que una secuencia, retomando las nociones centrales de la “semiosis social” [4]. Y así como la actividad de la secuencia genética está explicada por el Dogma Central, la semiosis genética podría ser explicada por lo que proponemos llamar un Dogma Periférico. Se trataría de un dogma, quizás no dogmático y abierto a múltiples correcciones, que asuma todo lo que se ha enumerado aquí en tanto periferia del centro del Dogma (DNA-RNA-proteínas), bajo la consideración del sistema DNA-RNA-proteínas-medio como un proceso semiótico y multidireccional.
Referencias:
1. Shannon C (1948) A Mathematical Theory of Communication Bell System Technical Journal 27: 379–423.
2. Jakobson R (1981) Ensayos de lingüística general Barcelona: Seix Barral.
3. de Saussure F (2007) Curso de lingüística general Buenos Aires: Losada.
4. Verón E (1998) La semiosis social Barcelona: Gedisa.
5. Barbero JM (1987) De los medios a las mediaciones. Comunicación, cultura y hegemonía Barcelona: Gustavo Gili.
6. Wiener N (1988) Cibernética y sociedad Buenos Aires: Sudamericana.
7. Sfez L (1995) Crítica de la comunicación Buenos Aires: Amorrortu.
8. Heims SJ (1991) The Cybernetics Group Cambridge: The MIT Press.
9. Crick F (1958) On protein synthesis Symposia of the Society of Experimental Biology 12: 138-63.
10. Crick F (1970). Central dogma of molecular biology Nature 227: 561–563.
11. Temin H, Mizutani S (1970) RNA-dependent DNA polymerase in virions of Rous sarcoma virus Nature 226: 1211–1213.
12. Shapiro JA (2009) Revisiting the Central Dogma in the 21st Century Annals of the NewYork Academy of Sciences 1178: 6–28.
13. Jackson D, Symons R, Berg P (1972) A Biochemical Method for Inserting New Genetic Information into SV40 DNA: Circular SV40 DNA Molecules Containing Lambda Phage Genes and the Galactose Operon of E. Coli Proceedings of the National Academy of Sciences of the United States of America 69: 2904.
14. Cohen S, Chang A, Boyer H, Helling R (1973) Construction of biologically functional bacterial plasmids in vitro Proceedings of the National Academy of Sciences of the United States of America 70: 3240–3244
15. Orgel L, Crick F (1980) Selfish DNA: The ultimate parasite Nature 284: 604–607.
16. Koonin E (2012) Does the central dogma still stand? Biology Direct, 7(1), 27-27.
17. De Lorenzo V (2014) From the selfish gene to selfish metabolism: revisiting the central dogma BioEssays, 36(3), 226-235
18. Bandyra K, Luisi B (2015) Central dogma alchemy RNA 21(4): 558-559.
19. Biro J (2014) Revisiting Crick’s Dogma and the Impossibility of Reverse Translation Theoretical Computer Science 1: 110.
20. Avery O, Macleod C, McCarty M (1944) Studies on the chemical nature of the substance inducing transformation of pneumococcal types: induction of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type III Journal of Experimental Medicine 79(2):137-58.
21. Jakobson R (1973) Essais de linguistique générale II : Rapports internes et externes du langage Paris: Les Éditions de Minuit.
22. Bell Enguix G, Jiménez López MD (2006) Código genético y lenguaje verbal Revista Española de Lingüística (RSEL) 36: 285-317.
El autor es investigador de CONICET
|
Revista QuímicaViva Número 2, año 14, Agosto 2015 quimicaviva@qb.fcen.uba.ar |